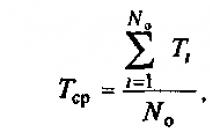Из истории изучения явления
Еще Чарльз Дарвин проводил исследования неблагоприятных результатов самоопыления, которые проявлялись в снижении темпов роста и жизненной активности перекрестноопыляющихся структур. Самоопыление, проводящееся на протяжении нескольких лет подряд, приводит к резкому уменьшению размера растений, снижению интенсивности их обмена веществ и падению их плодовитости.
Причиной резкого снижения жизнеспособности организмов при близкородственном скрещивании или самоопылении является высокая степень гомозиготности таких растений по многим генам. Подобная гомозиготность по рецессивным мутантным генам и является неблагоприятной для растений, подвергавшихся длительное время самоопылению, которое получило в генетике название инбридинга. В гетерозиготном состоянии вредное действие рецессивных генов (зачастую – летальных) не проявляется фенотипически. А в гомозиготном состоянии эти гены проявляются в фенотипе и оказывают неблагоприятное воздействие на организм.
Изучение генетики инбредных линий целого ряда растений позволило ученым разработать методы использования их в селекции для создания высокопродуктивных межвидовых гибридов. При правильном соединении таких инбредных линий удавалось при их скрещивании получить гибриды, которые по мощности своего развития значительно превышали исходные родительские формы.
Явление гетерозиса и его цитологические основы
Определение 1
Явление резкой вспышки гибридной мощности в первом поколении от скрещиваемых инбредных линий, а также сортов растений и пород животных, различающихся между собой по наследственным качествам, называется гетерозисом .
Иногда гетерозис называют еще «проявлением жизненной силы » или «гибридной силы ». В следующих гибридных поколениях гетерозис обычно угасает и через два-три поколения совсем исчезает.
Явление гетерозиса объясняется тем, что при скрещивании двух гомозиготных форм образуется гетерозиготный гибрид. В гетерозиготном состоянии летальные и сублетальные гены, как правило, - рецессивные. Поэтому они подавляются доминантными аллелями, и их вредное для организма действие не проявляются в фенотипе. Кроме того, в генотипе гибридных потомков могут сочетаться благоприятные доминантные аллели обоих родителей. В результате этого может наблюдаться явление взаимодействия неаллельных доминантных генов.
По данным современных биохимических наблюдений у гетерозисных форм наблюдается более широкий набор ферментов и их повышенная активность по сравнению с предками (родителями).
Ослабление гетерозиса в последующих поколениях объясняется переходом многих признаков (генов) снова в гомозиготное состояние. В восьмом поколении гибридов явление гетерозиса практически полностью исчезает.
У растений гетерозис можно закрепить, например, вегетативным размножением, удвоением числа хромосом или с помощью партеногенеза. Гетерозис может больше проявиться на одних признаках гибридной особи, не задевая других.
Значение гетерозиса
Явление гетерозиса широко применяют в сельском хозяйстве. Одним из первых растений, получение гетерозисных гибридов которого было поставлено на промышленную основу, стала кукуруза. Для получения гибридных семян сначала были созданы инбредные линии лучших сортов. Через $5-6$ лет инбридинга производят отбор лучших линий и испытывают их на наилучшее соединение. Линии с высокой комбинационной способностью, которые дают при гибридизации наибольший эффект гетерозиса, размножают и используют для массового получения гибридных семян.
Замечание 1
Применение гетерозиса позволяет многократно увеличить продуктивность сельского хозяйства. А это имеет важное значение для решения одной из важнейших глобальных проблем человечества – продовольственной проблемы.
Гетерозис - это свойство помесей и гибридов первого поколения (Fi) превосходить по биологическим и хозяйственно-полезным признакам исходные родительские формы.
Впервые явление гетерозиса было описано И. Кёльрейтером , работавшим в Петербургской академии наук, на примере межвидового гибрида, который он получил в 1760 г., скрестив два разных вида махорки . Этот растительный гибрид оказался стерильным наподобие мула и автор назвал его «первым растительным мулом».
Научный термин «гетерозис» появился гораздо позже. Его предложил американский исследователь Дж. Шелл в 1914 г. для обозначения мощности гибридов (эффекта скрещивания), и с тех пор он прочно вошел в научную литературу как синоним старого названия «гибридная сила».
В настоящее время твердо установлено, что гетерозис проявляется не только при разведении животных и птиц, но и при селекции растений, а также микроорганизмов. Следовательно, гетерозис - явление общебиологическое.
Каковы причины возникновения гетерозиса? На этот счет существует несколько точек зрения в виде отдельных самостоятельных гипотез.
Одна из первых изложена в фундаментальном труде Чарльза Дарвина «Действие перекрестного опыления и самоопыления в растительном мире», в котором он изложил вопросы гетерозиса («гибридной силы»).
Изучив опыт английских заводчиков по созданию новых пород сельскохозяйственных животных, Ч. Дарвин отметил, что родственное спаривание (инбридинг), которое применялось им для закрепления в потомстве желательных признаков выдающихся производителей, приводит, как и самоопыление у растений, к отрицательным последствиям - к депрессии, в то время как скрещивание повышает, как правило, жизнеспособность потомства за счет проявления эффекта скрещивания (гетерозиса).
Дарвин предположил, что в основе этих двух явлений - инбредной депрессии и гетерозиса - лежит одна и та же причина - степень различия половых элементов, объединяющихся в процессе оплодотворения.
Чем больше родительские формы, а следовательно, и их половые клетки различаются по своим биологическим особенностям, тем сильнее в потомстве проявляется гетерозис, и наоборот, отсутствие таких различий при близкородственном длительном спаривании приводит к нибредной депресии.
Исходя из этих соображений, Ч. Дарвин провозгласил «великий закон природы», по которому, сточки зрения эволюции вида, скрещивание всегда полезно, а родственное спаривание (инбридинг - у животных, самоопыление - у растений) - вредно.
Гипотеза доминирования (Джонс, 1972). В основе этой гипотезы лежит идея благоприятного действия доминантных генов - гетерозис проявляется в результате взаимодействия при скрещивании благоприятных доминантных факторов, имеющихся у исходных родительских форм.
Предполагается, что при скрещивании происходит сочетание благоприятно действующих неаллельных доминантных генов и одновременное подавление ими действия различных вредных рецессивных аллелей, которые у разных линий, а тем более пород, находятся в разных локусах. При скрещивани доминантные аллели, внесенные одним родителем (линией), могут перекрывать рецессивные аллели, полученные гибридом от другой родительской формы (линии).
Проявление гетерозиса возможно также и за счет явления эпистаза, когда отдельные неаллельные гены (эпистатический ген) подавляют не только «свои» рецессивные, но и «чужие» доминантные гены (гипостатический ген).
Гипотеза доминирования общепризнана, однако, она не объясняет полностью все вопросы, возникающие в связи с проявлением гетерозиса. Так, если исходить из названной гипотезы, то теоретически следует ожидать, что при полигибридном скрещивании гетерозигота Аа будет в той или иной степени приближаться по продуктивности к гомозиготе АА - приближаться, но не превосходить ее.
Однако в практике давно установлено, что гетерозигота может превосходить по мощности не только рецессивную родительскую форму, но и доминантную, то есть обоих своих родителей. Это явление получило в генетике даже специальное название - сверхдоминирование, или моногибридный гетерозис.
Гипотеза гетерозиготности (сверхдоминирование). С точки зрения гипотезы гетерозиготности проявление гетерозиса объясняется, говоря современным языком, разнокачественностью членов одной и той же пары аллелей у гибридных организмов как результат скрещивания различающихся исходных родительских форм. Соединение при гибридизации разнокачественных гамет родителей уже само по себе стимулирует более быстрый рост гетерозиготных гибридов, их лучшее развитие и т. д. В результате гибрид по мощности превосходит исходные гомозиготные родительские формы как рецессивную, так и доминантную, что и обусловливает эффект сверхдоминирования. В то же время гомозиготность родителей оказывает угнетающее действие на жизнеспособность потомства, что выражается формулой Аа>АА>аа.
Предполагается, что у гетерозиготы оба аллеля одного локуса выполняют различные функции, взаимно дополняя друг друга в биохимическом процессе. При этом эффект гетерозиса будет тем выше, чем в большей степени аллели каждого локуса различаются функционально между собой, чем больше они дополняют друг друга.
Причины гетерозиса, указанные в этих гипотезах, возможно, действуют одновременно, однако их, по-видимому, не достаточно для всестороннего объяснения механизма возникновения гетерозиса как общебиологического явления. По этому поводу проф. М. В. Лобашов (1969) писал: «Трудно предположить, чтобы в основе столь сложного явления, как гетерозис, лежал единственно генетический механизм». Что же касается понимания самого механизма генного взаимодействия при гетерозисе, то по современным воззрениям различие между обеими гипотезами незначительно или его совсем нет.
Гипотеза генетического баланса (академик ВАСХНИЛ Н. В. Турбин, 1961. С точки зрения этой гипотезы явление гетерозиса нельзя объяснить действием одной какой-либо генетической причины - это суммарный эффект.
Гипотеза генетического баланса , принимая в обеих чертах отдельные положения ранее высказанных гипотез, больше внимания, однако, уделяет взаимовлиянию неаллельных генов, физическим и биохимическим факторам, а также внешней среде вообще, условиям выращивания гибридов в частности . Особое внимание придается цитоплазматическому влиянию. Предполагается, что плазматические различия между гаметами должны стимулировать жизненные процессы у гибридного организма. Сбалансированность систем генов делает популяции наиболее приспособительными и продуктивными в конкретных условиях среды.
Следует отметить, что в последние годы все большее значение приобретает также биохимическая теория гетерозиса , согласно которой скрещивание приводит к увеличению гетерозиготности по мутациям, регулирующим синтез белка - отсюда проявление гетерозиса происходит за счет обогащения биохимических процессов в клетках и тканях гибридного организма.
Значение изложенных выше гипотез неоспоримо, однако ни одна из них не может быть пока признанной в качестве общепринятой теории гетерозиса. Возможно, прав известный генетик Ф. Хатт, в своем высказывании: «Гетерозис все еще представляет одну из самых больших загадок генетики».
Что же представляет собой гетерозис как генетическое явление? Каковы формы проявления гетерозиса?
Гетерозис - это совокупность явлений, связанных с повышенной жизнеспособностью помесей (гибридов), которая, как установлено, проявляется уже на ранних стадиях развития (онтогенеза) . У эмбрионов гибридных кур, например, еще в эмбриональном развитии усиливаются обменные процессы, ускоряется их развитие, в результате чего вывод и качество суточного молодняка бывают выше по сравнению с теми же показателями линейных кур (Злочевская, 1968).
Гетерозис - явление неустойчивое (кратковременное), оно наиболее ярко (четко) проявляется лишь в первом поколении (F 1) скрещивания. Помесные (гибридные) животные при их дальнейшем разведении не дают подобных себе гетерозисных потомков, онине остаются «константными» по гетерозису. Поэтому их не оставляют на племя, а реализуют на мясо. Следовательно, гетерозис нельзя закрепить наследственно, его нужно всякий раз получать заново.
В отдельных случаях гетерозис можно поддерживать на относительно высоком уровне и в последующих поколениях, но в таких случаях используются специальные методы - переменное скрещивание и др. Считается, что угасание гетерозиса в последующих поколениях гибридов - результат рекомбинационных утерь .
Формы проявления гетерозиса бывают различными. Обычно при скрещивании двух пород (А и В) уровень продуктивности помесного (АВ) потомства равен средней показателей продуктивности исходных пород В таких случаях говорят о гипотетическом (вероятном) гетерозисе.
![]()
Нередко продуктивность помесных (F 1) животных оказывается значительно выше средней продуктивности родителей, а иногда она превышает показатели лучшей из родительских форм - абсолютный (истинный) гетерозис.
![]()
В других случаях, однако, продуктивность помесей превышает показатели лишь одного из родителей, худшего - относительный гетерозис:
![]()
где: Пг - признак гибрида; Пл - признак лучшей породы; Пм - признак материнской породы; По - признак отцовской породы.
Конечно, из чисто практических соображений эффект гетерозиса представляет наибольший интерес лишь в том случае, когда гибридное потомство превышает по своей общей хозяйственно полезной ценности лучшего из родителей. Только в таких случаях скрещивание имеет экономический смысл. Поэтому селекционеры-практики под гетерозисом понимают свойство гибридов (F 1) превосходить по определенным признакам лучшую из родительских форм.
Некоторые ученые, учитывая специфику форм проявления гетерозиса, выделяют самостоятельные его типы:
репродуктивный гетерозис - более высокая общая продуктивность животных, связанная с повышением плодовитости (фертильности) и более мощным развитием их репродуктивных органов;
соматический гетерозис - более сильное развитие вегетативных частей (у растений), органов и частей тела (у животных);
адаптивный гетерозис - повышенная жизнеспособность животных, их лучшая приспособляемость.
У мулов, например, сильно выражен соматический гетерозис, то есть большая живая масса; выше тяговое усилие; повышенное долголетие; особая выносливость; но в то же время репродуктивная система недоразвита. Как правило, они бесплодны. Сказанное выше - это пример частного гетерозиса (мощное развитие касается не всего организма животного, а лишь его отдельных признаков) в отличие от общего гетерозиса, когда имеет место развитие общей массы тела животного, повышение метаболических процессов в организме в целом, что обеспечивает повышение его продуктивности (Красота, 1979).
Следует отметить, что гетерозис проявляется у помесей и гибридов - межвидовых, межпородных, межлинейных - по ограниченному числу признаков. Он никогда не проявляется по сумме всех родительских признаков. Помеси (гибриды) превосходят своих родителей не по всем показателям продуктивности, не по всем признакам, не по их сумме, а лишь частично, по отдельным признакам (или группе признаков) или даже по отдельно взятому признаку.
Метисные куры первого поколения, полученные от скрещивания петухов мясных пород с курами яйценоских (легких) пород, могут превосходить исходные родительские формы по яйценоскости, но по живой массе они занимают промежуточное положение.
Отсюда под гетерозисом следует понимать превосходство потомства - помесей или гибридов - над родительскими формами не по всем, а лишь по определенным, конкретным признакам.
Литература
1. Вавилов Н. И. Центры происхождения культурных растений.- Тр. по прикладной ботанике и селекции, 1926, т. XVI, с. 5-138.
2. Гершензон С. М. Основы современной генетики.- Киев, 1979.- 506 с.
3. Лобашев М. Е., Ватти К. В., Тихомирова М. М. Генетика с основами селекции.- М., 1979.- 304 с.
4. Рокицкий П. Ф. Некоторые этапы развития генетики животных в СССР и ее связи с селекцией.- В сб.: Генетические основы селекции животных. М., 1969, с. 9-25.
Знакомы ли вы с таким понятием, как эффект гетерозиса? В нашей статье мы рассмотрим его причины, области применения и значение для человека.
Системы гибридизации
С целью совершенствования существующих и выведения новых сортов растений, пород животных и штаммов бактерий человек два основных метода селекции. Это искусственный отбор и гибридизация. Сначала он отбирает из исходного материала особей с наиболее ценными свойствами. Это могут быть показатели яйценоскости кур, количества крахмала в семенах, мышечной массы крупного рогатого скота, размера плодов. Для увеличения эффективности учитывают генотип и состояние аллелей исходных форм.
Результат во многом зависит и от системы скрещиваний - гибридизации. Суть этого метода заключается в соединении генетического материала половых или соматических клеток.
Родственное скрещивание
Существует несколько типов гибридизации. Родственное скрещивание, или инбридинг является результатом соединения генетического аппарата особей одного вида. Оно возможно среди самоопыляемых растений и гермафродитных животных, способных к самооплодотворению.
Каковы последствия такой гибридизации? В каждом последующем поколении повышается количество гомозиготных особей. Их фенотипические признаки постепенно ослабляются, что приводит к вырождению линии.

Отдаленная гибридизация
Знаете ли вы, кто является исходными формами мула? Это осел и кобыла. А если соединить генотипы одно- и двугорбого верблюда, то получится гораздо более сильный и выносливый гибрид. Все это примеры отдаленной гибридизации.
При этой системе скрещивания возникает только одна проблема. Межвидовые гибриды бесплодны. Исправить это возможно только путем кратного увеличения хромосомного набора.

Неродственное скрещивание
Явление гетерозиса связано с гибридизацией разных сортов, пород и линий, которые относятся к одному виду. Такое скрещивание называется неродственным, или аутбридингом.
Оно имеет обратный эффект. С каждым последующим поколением возрастает гомозиготность. При этом мутации, имеющие негативные последствия, не проявляются. Они переходят в гетерозиготное состояние и не оказывают влияния на жизнеспособность видов. Поэтому этот вид гибридизации является основой селекции.

Явление гетерозиса
Ученые давно заметили, что соединяя генотипы неродственных особей одного вида, полученные при этом гибриды отличаются повышенной жизнеспособностью. Они более плодовиты, устойчивы к действию болезнетворных микроорганизмов. Такое явление называют гетерозисом. неродственного скрещивания. В переводе с греческого данный термин означает "перевоплощение".
Эффект гетерозиса обусловлен тем, что летальные и сублетальные аллели могут переходить в гетерозиготное состояние. Благодаря этому их негативные последствия внешне не проявляются. При этом генотип гибридов первого поколения часто содержит доминантные аллели благоприятных признаков.

Эффект гетерозиса
Гибридная сила является понятием противоположным инбридингу, или близкородственному скрещиванию, результатом которого является фенотипическая депрессия особей.
Гетерозис проявляется не только в форме повышения гетерозиготности. В таком состоянии формируется несколько видов ферментов. Их совокупное действие также повышает гибридную силу потомков.
Впервые гетерозис был открыт еще в 19-м веке немецким ботаником Кельрейтером. Ему принадлежат первые опыты по искусственной гибридизации растений. Причем произошло это еще до того, как Грегори Мендель сформулировал свои знаменитые законы о наследственной изменчивости. В начале 20-го века был подробно описан гетерозис кукурузы. Урожай ее гибридных образцов во много раз превышал этот показатель у обычных форм.
Ученые различают несколько форм гетерозиса. Но так или иначе все они проявляются в виде гибридной силы. При репродуктивной форме наблюдается повышение плодородности и урожайности, поэтому она присуща растениям. Гибридная сила проявляется и в способности организмов противостоять неблагоприятным условиям среды. Такая форма называется приспособительной, или адаптивной. Соматический гетерозис приводит к увеличению линейных размеров и массы неродственных гибридов, полученных в первом поколении.
Область применения
Поскольку гетерозис - это явление, при котором значительно повышаются производительные качества растений и животных, оно широко применяется в сельском хозяйстве.
В каких областях возможно его практическое применение? Прежде всего, в птицеводстве. Наиболее эффективными являются две линии. Это выведение яичных и бройлерных пород. Гетерозис используют в таких направлениях животноводства, как рыболовство, овцеводство, мясное скотоводство, верблюдоводство.
Ученые установили, что гибридная сила может быть последствием и межвидового скрещивания. Мулы гораздо более выносливее и сильнее по сравнению со своими родительскими формами. В начале прошлого века в Соединенных Штатах была выведена известная мясная порода Санта Гертруда. Это гибрид зебу с шортгорнами - коровами мясного направления. Результат был просто потрясающим. Вес взрослых особей полученных гибридов приближается к 700 кг.
Еще одним примером проявления гибридной силы является порода овец архаромеринос. Она была выведена в Казахстане в середине 20-го века в результате скрещивания тонкорунных овец с горными баранами архарами. Полученные гибриды приобрели ценные признаки сразу в нескольких направлениях: настриг шерсти, скороспелость, плодовитость, убойный выход, приспособленность к горным условиям, сильный иммунитет.
Гетерозис - это метод, которые широко используется и в растениеводстве. Этот прием позволяет повысить урожайность отдельных сортов до 30 %. Важными аспектами являются получение гибридных семян. Гетерозис при этом позволяет сделать этот процесс экономически рентабельным.
Перспективным направлением селекции является закрепление эффекта гетерозиса в генотипе растений, способных к вегетативному размножению. Чаще всего для этого используют плодово-ягодные и клубневые культуры, которые получены из гибридных семян.
Лидирующее место среди гетерозисных культур по-прежнему занимает кукуруза. Выведены гибридные сорта этого растения, которые отличаются засухоустойчивостью, высоким иммунитетом, хорошими показателями продолжительности сохранности семян. Широко распространения также получили гибриды сахарной свеклы, сорго, пшеницы.

Существенные недостатки
Явление гетерозиса зависит от степени родства между родительскими особями: чем более отдалёнными родственниками являются родительские особи, тем в большей степени проявляется эффект гетерозиса у гибридов первого поколения.
В чем главная проблема гибридной силы? Его проявление наблюдается только в первом поколении. Во втором оно ослабевает, а в последующих и вовсе сходит на нет. В настоящее время селекционеры пытаются найти пути выхода из данной ситуации. Один из них - переменное скрещивание. Областью его применения является товарное животноводство. В ходе этого процесса сначала получают гибридов двух разных пород. Потом полученная особь также скрещивается. Но при этом используется особь уже совсем другой породы.
Итак, эффект гетерозиса обусловлен повышением гетерозиготности организмов и переходом летальных аллелей в рецессивное состояние. Фенотипически его эффект проявляется в увеличении количественных и качественных показателей гибридов первого поколения в сравнении с родительскими формами.
Гетерозис , или гибридная мощность (сила) – это явление превосходства гибридов 1-го поколения по сравнению с исходными родительскими формами. Оно проявляется по многим признакам при скрещивании разных видов, рас, пород животных и сортов растений, а также инбредных линий.
Еще в середине XVIII в И. Кельрейтер , академик Российской академии, знаменитый ботаник, обратил внимание на тот факт, что в отдельных случаях при скрещивании растений гибриды первого поколения значительно мощнее своих родительских форм. Затем Ч. Дарвин сделал заключение, что гибридизация во многих случаях сопровождается более мощным развитием гибридных организмов.
Термин «гетерозис » ввел в 1914 г . американский генетик и кукурузовод Дж. Шелл .
Хотя эффект гетерозиса известен с древнейших времен, его природа до начала XX в. была неясна. Глубокий научный анализ явления гетерозиса стал возможен только после открытия основных генетических закономерностей.
Межлинейные гибриды кукурузы . В начале XX века Г. Шелл показал, что при скрещивании некоторых инбредных линий кукурузы получаются дает гибридные растения, более урожайные по зерну и вегетативной массе, чем исходные линии и сорта.
Сейчас посев гибридными семенами стал основным приемом производства кукурузы . Для получения гибридных семян сначала создают большое количество инбредных линий из лучших сортов, отвечающих требованиям данного климатического района. Инбредная линия создается в течение 5 – 7 лет путем самоопыления . При отборе линий оцениваются качества, которые необходимо получить у будущего гибридного потомства. Значительная часть линий (около 99%) бракуется из-за тех или иных отрицательных свойств.
Создание большого количества инбредных линий – необходимый этап работы для получения гетерозисных форм. Особи в пределах линии имеют сходные генотипы и является практически гомозиготными. Поэтому при скрещивании таких линий образуются одинаковые по генотипу гетерозиготные гибриды .
Полученные таким образом межлинейные гибриды первого поколения оценивают по эффекту гетерозиса, отбирают линии, дающие лучшие комбинации, и затем размножают их в больших масштабах для производства гибридных семян . Чем больше создано инбредных линий, тем вернее и скорее можно отыскать лучшие гибридные комбинации с необходимым сочетанием свойств. Чтобы найти пару линий , дающих при скрещивании высокий эффект гетерозиса , необходимо проверить несколько тысяч гибридных комбинаций .
При получении гибридных семян для производственных целей исходные линии , дающие при скрещивании наибольший эффект гетерозиса, высевают рядами, чередуя материнские и отцовские формы . Для обеспечения опыления между ними разработана схема производства гибридных семян с использованием цитоплазматической мужской стерильности , что позволило значительно сократить затраты труда на удаление метелок с растений материнской линии . Так получают простые межлинейные гибриды кукурузы. Этот метод в принципе является общим для семеноводства гибридов различных перекрестно опыляющихся растений. Бывают также тройные гибриды (рис. 226).
В настоящее время в практике сельского хозяйства простые межлинейные гибриды кукурузы не используются, так как затраты на получение таких семян не окупаются. Теперь широко внедряется в практику посев семян двойных межлинейных гибридов . Последние получают путем скрещивания двух простых гибридов, проявляющих гетерозис (рис. 147).
Тройные идвойные гибриды получают в два этапа: 1) получение простого гибрида; 2) использование простого гибрида в качестве материнской формы для тройного или двойного, в качестве отцовских форм которых выступают линия и простой гибрид соответственно.
Подбор простых гибридов для получения наиболее продуктивных двойных гибридов является важным этапом селекции. Лучшие результаты дает скрещивание инбредных линий , происходящих из различных сортов . Так, например, если один простой гибрид получен от скрещивания инбредных линий двух сортов – А × В , а другой – от скрещивания линий других сортов – C × D , то двойной гибрид (А × В ) × ( C × D ) дает гетерозис чаще, чем если бы двойной гибрид был получен от скрещивания простых гибридов, каждый из которых происходит от линий одного сорта. Совершенно ясно, что в большинстве случаев инбредные линии всегда будут иметь более низкие показатели, чем сорта. О наличии гетерозиса следует говорить лишь в том случае, когда межлинейный гибрид превосходит не только исходные линии , но и сорта или породы , от которых произошли эти линии.
Применение цитоплазматической мужской стерильности. Возникает вопрос, как получить гибридные семена, например, у кукурузы, сахарной свеклы, риса, томатов, если в пределах одного растения или даже одного цветка расположены женские и мужские элементы системы размножения и всегда присутствует возможность самоопыления. В этих случаях избежать процесса самоопыления можно только двумя путями: на материнских формах удалить вручную мужские элементы цветка, продуцирующие пыльцу; сделать мужские соцветия стерильными . Первый путь очень трудоемок, поэтому генетики начали поиск систем, определяющих мужскую стерильность растений.
В 1929 г. ученик Н. И. Вавилова отечественный селекционер и генетик Михаил Иванович Хаджинов нашел в посевах кукурузы растения с мужской стерильностью, которые ничем не отличались от нормальных, только были полностью стерильными, т. е. не продуцировали пыльцу. Это явление – цитоплазматическая мужская стерильность (ЦМС) – было изучено и широко использовано для получения гибридных семян у кукурузы, а затем и у многих других видов.
Схема использования ЦМС в селекции разработана в 30-х годах М. Родсом . Было установлено, что только взаимодействие особого типа цитоплазмы (S) и рецессивных генов ядра (rf) обусловливает мужскую стерильность (рис. 227, 228).
В практике используют лишь гибридные семена первого поколения от скрещивания двух линий, простого гибрида и линии или двух простых гибридов. Второе и последующие поколения в производственных посевах не используются, так как гибриды расщепляются на исходные формы и эффект гетерозиса исчезает. В связи с этим при использовании гетерозиса у растений организовано семеноводство в специальных хозяйствах, где получают только семена первого поколения и продают их хозяйствам, фермерам и т. д. Так как урожайность гетерозисных гибридов значительно (на 20-30%) выше исходных сортов, то затраты на семеноводство гибридных семян с лихвой окупаются. Внедрение гетерозисных гибридов кукурузы, по оценке американских специалистов, принесло чистый доход, в сотни миллиардов долларов. Гетерозис используется также при выращивании сахарной свеклы, риса, томатов и других видов.
Аналогичным образом получают гибриды и у животных. В настоящее время в птицеводстве и в свиноводстве широко используется скрещивание инбредных линий, происходящих из одной или разных пород .
Факторы , влияющие на гетерозис. Проявление гетерозиса у гибрида зависит:
1) от генотипа;
2) от свойств цитоплазмы , в результате чего реципрокные скрещивания дают разный эффект . Например, от скрещивания ♀ лошадь × ♂ осел получается высокогетерозисный гибрид мул – долговечный, выносливый и сильный. Реципрокная комбинация дает лошака , у которого гетерозис полностью отсутствует .
3) от стадии онтогенеза. Гетерозис в онтогенезе реализуется неравномерно . На одних стадиях онтогенеза проявляется гетерозис по одним признакам, на других – по другим. Так, в раннем возрасте у одного и того же гибрида может наблюдаться гетерозис в отношении скорости роста отдельных частей организма и повышенной устойчивости к заболеваниям, но его может не быть , например, в отношении устойчивости к неблагоприятной температуре . Гетерозис по этому свойству может проявиться позднее .
4) Сильное влияние на проявление гетерозиса оказывают также факторы среды .
Возможные механизмы гетерозиса. В настоящее время имеются 4 гипотезы , объясняющие возникновение гетерозиса:
1. Гипотеза гетерозиготности гибридов Дж. Шелла и Е. Иста (1908 г.). Как уже говорилось, при скрещивании гомозиготных инбредных линий гибриды первого поколения гетерозиготны по многим генам . При этом действие вредных рецессивных мутантных аллелей подавляется доминантными аллелями обоих родителей.
Схематически это можно представить так: одна инбредная линия в гомозиготном состоянии имеет рецессивную аллель одного гена (ааВВ) , а вторая – другого гена (ААbb) . Каждая из этих рецессивных аллелей в гомозиготном состоянии определяет какую-нибудь недостаточность, которая снижает жизнеспособность инбредной линии.
При скрещивании линий aaBB × AAbb у гибрида объединяются доминантные аллели обоих генов (AaBb ) . Гибриды F 1 проявят при этом по указанным генам не только гетерозис , но и единообразие .
ВF 2 число особей с двумя доминантными генами в гетерозиготном состоянии (AaBb) будет лишь 4 / 16 , поэтому гетерозисными оказываются не все особи. В дальнейших поколениях число гетерозигот сокращается, а число гомозигот увеличивается . В силу этих причин гетерозис в последующих поколениях затухает .
Серьезным препятствием для этой гипотезы является тот факт, что далеко не все межлинейные гетерозиготные гибриды проявляют гетерозис , т. е. гетерозиготность не всегда связана с гетерозисом.
2. Гипотеза доминирования Д. Джонса (1918 г.) исходит из того факта, что доминантные аллели дикого типа чаще, чем рецессивные, оказывают благоприятное действие независимо от того, в каком состоянии - гомозиготном или гетерозиготном – они находятся. Поэтому подбор в гибридной комбинации доминантных аллелей может скорее обеспечить гетерозис. Иначе говоря, эта гипотеза исходит из представления о простом суммировании эффекта доминантных аллелей с комплементарным действием .
Проиллюстрируем это следующим примером :
P: AAbbCCdd × aaBBccDD
Если у скрещиваемых форм имеется всего по два доминантных благоприятно действующих гена, то у гибрида их четыре, независимо от того, в гомозиготном или гетерозиготном состоянии они находятся. Это, по мнению сторонников этой гипотезы, и определяет гетерозис гибрида, т. е. его преимущества перед исходными формами.
И эта гипотеза вступает в противоречие с рядом фактов. Так, при инбридинге идет гомозиготизация . Следовательно, согласно этой гипотезе, следовало ожидать появления в результате инбридинга гетерозисных форм , имеющих набор доминантных генов в гомозиготном состоянии, однако это не имеет места.
3. Гипотеза сверхдоминирования Шелла и Иста исходит из того, что гетерозиготное состояние имеет превосходство над гомозиготным (АА < Аа > аа). Предполагается, что сочетание в гетерозиготе аллелейдикого типа и мутантного каким-то образом усиливает действие доминантного гена и в связи с этим вызывает максимальное накопление специфических веществ , синтез которых контролируется этим геном.
4. Гипотеза компенсационного комплекса генов В. А. Струнникова . Ее суть сводится к тому, что при возникновении мутаций сильно понижающих жизнеспособность и продуктивность организма, так называемых полулеталей , при отборе у гомозигот формируется компенсационный комплекс генов , в значительной степени нейтрализующий вредное действие мутаций. Если затем такую мутантную форму скрестить с нормальной без мутации и тем самым перевести мутацию в гетерозиготное состояние , т. е. нейтрализовать ее действие нормальным аллелем, то сложившийся по отношению к мутации компенсационный комплекс в гибридном организме будет «работать» на гетерозис .
Пути закрепления гетерозиса. Основной задачей использования гетерозиса в селекции является закрепление его , т. е. сохранение эффекта гетерозиса в процессе воспроизведения гибрида . Решение этой задачи возможно следующими путями.
1. Закрепить гетерозис удается там, где возможно вегетативное размножение гибридов – клубнями, луковицами, черенками. Так сохраняется положительный эффект гетерозиса у картофеля.
2. Важным подходом к закреплению гетерозиса является кратное увеличение наборов хромосом – полиплоидия .
3. В естественных условиях гетерозис закрепляется при возникновении полиморфизма по инверсиям .
4. У животных, для которых все эти пути закрепления гетерозиса исключены, используют переменное скрещивание , т. е. регулярное скрещивание гибридов попеременно с одной и с другой исходными формами.
5. Аналогично опытам Гердона возможна пересадка ядер гетерозисных животных в энуклеированные яйцеклетки, т. е. клонирование .
Установите соответствие между методами и областями науки и производства, в которых эти методы используются: к каждой позиции, данной в первом столбце, подберите соответствующую позицию из второго столбца.
Запишите в таблицу выбранные цифры под соответствующими буквами.
| A | Б | В | Г | Д | Е |
Пояснение.
Биотехнология - это производство необходимых человеку продуктов и материалов с помощью живых организмов, культивируемых клеток и биологических процессов.
Селекция: получение полиплоидов; испытание по потомству; гетерозис. Биотехнология: метод культуры клеток и тканей; использование дрожжей для производства белков и витаминов; метод рекомбинантных плазмид.
Ответ: 122211.
Примечание .
Плазмиды – мелкие кольцевые молекулы ДНК, присутствующие в клетках бактерий. Они содержат дополнительную генетическую информацию, способны автономно, независимо от ДНК хромосом, реплицироваться; некоторые плазмиды обладают способностью встраиваться в хромосому бактерии и выходить из нее; некоторые могут переходить из одной клетки в другую. В генетической инженерии наиболее широко используется три типа плазмид F, P и Col. Метод создания рекомбинантных плазмид был разработан П.Бергом в 1972г. Ими была создана рекомбинантная плазмида, содержащая галактозный оперон E.coli. В плазмиду могут быть включены природные или синтезированные гены. После проникновения в клетку бактерии рекомбинантная плазмида может функционировать и размножаться автономно, либо включаться в ДНК хромосомы бактерии. Таким методом в клетки бактерии были введены гены человека и созданы штаммы бактерий-суперпродуценто соматостатина, интерферона, инсулина, гормоны роста человека, быка, глобин животных и человека.
Разработка биотехнологии производства интерферона – сложный процесс, требующий строгой регламентации действий на многочисленных этапах. Рассмотрим получение интерферона с помощью технологии рекомбинантной ДНК . Рекомбинантную молекулу ДНК получают встраиванием определенных генов в ДНК. С помощью ферменто-рестриктаз «разрезают» участки исходной ДНК и выделяют нужные гены. Другой фермент – лигаза – «вшивает» гены в новую ДНК. Микроорганизмы с рекомбинантной ДНК при их выращивании производят нужный продукт.
Вначале выделенную из крови доноров и находящуюся в культуре суспензию лейкоцитарных клеток обрабатывают вирусом, оказывающим индуцирующий эффект на биосинтез интерферона. В дальнейшем из лейкоцитов получают иРНК, программирующую биосинтез интерферона. Даже в индуцированных вирусом Сендай лейкоцитах иРНК содержится не более 0,1 % (Смородинцев А.А., 1985).
С помощью фермента обратной трансриптазы (ревертазы) на полинуклеотидной основе иРНК синтезируют комплементарную ей одноцепочечную копию ДНК (кДНК). Этому этапу предшествует синтез дезоксирибонуклеотида – затравки, состоящей из 32 мононуклеотидов, которая при гибридизации взаимодействует с соответствующим комплементарным участком выделенной из лейкоцитов иРНК и в дальнейшем выступает в качестве стартовой отметки, от которой начинается РНК-зависимый синтез одной из цепей ДНК (кДНК).
На следующем этапе на отделенной от гибридной структуры ДНК –РНК одноцепочечной кДНК осуществляется биосинтез второй комплементарной цепи ДНК. Чтобы обеспечить в синтезированной к ДНК комплементарность липких концов, к ним присоединяются линкеры (переходники). Они являются синтезированными химическим путем короткими участками ДНК, имеющими разные липкие концы. Обработка рестрикционной эндонуклеазой концов кДНК, а также подобранной плазмиды вектора. Которая в результате ферментативного гидролиза расщепляется рестриктазой с образованием линейной молекулы ДНК с липкими концами, позволяет соединить кДНК с плазмидой и благодаря липким концам и с помощью ДНК-лигазы образовать кольцевидную рекомбинантную плазмиду с синтезированной кДНК, в которой находится ген, кодирующий биосинтез а-интерферона.
Затем рекомбинантную плазмиду необходимо ввести в бактериальную клетку. Следующий этап – поиск бактериальной клетки, содержащей ген интерферона. По такому признаку, как способность к гибридизации, идентифицируют бактерии, содержащие рекомбинантные плазмиды с включенным в них геном, кодирующим синтез интерферона. Эти рекомбинантные плазмиды из бактерий выделяют и с помощью рестриктаз получают гены интерферона. Эукариотический интерфероновый ген в бактериальной клетке кодирует синтез «сырого» интерферона, для превращения которого зрелый интерферон в эукариотических клетках нет необходимых условий. Для преодоления этого препятствия эукариотический ген в условиях in vitro перестраивают, удаляя соответствующей рестриктазой ту часть нуклеотидов, которые кодируют информацию, не входящую в молекулу функционального интерферона. При этом в ходе рестриктазной реакции получается «перебор». Одновременно удаляется триплет, кодирующий синтез первой аминокислоты в полипептидной цепи интерферона. Этот, а также предшествующий ему инициирующий биосинтез полипептидной цепи кодоны синтезируют химическим путем и присоединяют к гену интерферона. Созданный в результате сложных манимуляций ген переносится в плазмиду, где он совмещается с бактериальным промотором, а затем вводится в бактериальную клетку-хозяин. Таким сложным многоэтапным путем создан штамм-продуцент E.coli. В 1 л бактериальной суспензии, содержащей около 1011 клеток, концентрация а-интерферона достигает 5 мг, что в 5 тыс. раз больше того количества, которое можно получить из 1 л донорской крови.