Leucemia (leucemia) este o boală tumorală a țesutului hematopoietic. Ele sunt caracterizate prin proliferarea sistemică a celulelor hematopoietice imature în diferite organe și țesuturi. Diferențele interspecifice și încrucișate. Leucoza este mai frecventă la bovine decât la oi, cai și porci. În aceleași condiții de mediu, nu există diferențe în incidența leucemiei între rase. Influența producătorilor și a liniilor. La toate rasele, s-au evidențiat diferențe mari în incidența leucemiei între derivați. În cadrul fermelor individuale, au fost identificate diferențe între liniile de incidență a leucemiei. Influența familiilor și a mamelor. Există diferențe mari între familii în ceea ce privește incidența leucemiei. Există familii fără leucemie și cele cu morbiditate. Mamele cu leucemie au născut de 2 ori mai multe fiice bolnave decât fiice sănătoase. Incidența îmbolnăvirilor la nepoate depinde într-o măsură mai mică de starea de sănătate a bunicilor. Incidența leucemiei până la vârsta de 3 ani este mai mică decât incidența la vârste mai înaintate. Moștenirea rezistenței și susceptibilității la leucemie. Rezistența la leucemie este determinată de mulți loci genici. La pui, susceptibilitatea la infecția cu virusul leucemiei domină asupra rezistenței și este o trăsătură monogenă. Se crede că leucemia este cauzată de virusul leucemiei care conține ARN k.r.s. Yashur. Acut boala virala artiodactilii. Zebui sunt relativ rezistenți la boală. boala lui Marek - boala infectioasa păsări (agentul cauzal este un virus care conține ADN), caracterizat prin proliferarea țesutului limforreticular în organele interne, piele, mușchi și deteriorarea trunchiurilor nervoase periferice. Unele rase de pui variază în ceea ce privește rezistența la boala Marek. Rezistența la boală domină asupra susceptibilității. Pseudoplaga păsărilor - caracterizată prin pneumonie, encefalită. Păsările lipsite de limfocitele T sunt foarte susceptibile la boală: Scrapie (scrapie) este o boală infecțioasă care se dezvoltă lent, care duce la modificări distrofice ale sistemului nervos central. Cauzat de un agent asemănător virusului, a cărui natură nu este clară. Un semn caracteristic al bolii este mâncărimea. Alela dominantă controlează susceptibilitatea la scrapie, în timp ce alela recesivă controlează rezistența. Controlul genetic al rezistenței la scrapie depinde de tulpina patogenului. Mixomatoza de iepure. O boală virală acută caracterizată prin conjunctivită sero-purulentă și formarea de tumori la cap, anus și organe genitale externe
Leucemia (leucemia, leucemia) sunt boli tumorale ale țesutului hematopoietic. Ele sunt caracterizate în principal prin proliferarea sistemică a celulelor hematopoietice imature în diferite organe și țesuturi.
Diferențele interspecifice și încrucișate. Leucemia este mai frecventă la mare bovine decat 4 oi, cai si porci. Melanoamele maligne sunt raportate mai frecvent la cai.
Dintre cele 34 de rase de animale din țară, animalele cele mai susceptibile la leucemie sunt rasele roșii (letonă brună, daneză roșie, lituaniană roșie etc.) și rasele alb-negru (Nakhameon, 1989). Leucemia apare rar la rasele de origine elvețiană (Kostroma, Lebedinskaya, Brown Carpathian). Printre rasele din Germania de Nord, leucemia-1* a fost înregistrată de 8 ori mai des decât la rasele bavareze. Se presupune că rasa roșie Gorbatov, bovinele roșii Megrelian, Yakut, Suksun și Bushuevsky sunt relativ rezistente la leucemie (Dmitriev și colab., 1986). Rata de infectare a raselor de lapte este semnificativ mai mare decât cea a raselor de carne. Cu toate acestea, unii cercetători cred că în aceleași condiții de mediu nu există diferențe în incidența leucemiei între rase.
Influența producătorilor și a liniilor. La toate rasele, s-au evidențiat diferențe mari în ceea ce privește incidența leucemiei la descendenți între producători. Astfel, frecvența fiicelor afectate ale unor tauri este de 0-5%, altele - 20-50% și mai mare. Influența taților asupra rezistenței fiicelor la leucemie este de 15%, iar în unele efective ponderea de influență este de 3-5%. Încrucișarea taților rezistenți cu mame sănătoase și leucemice a dus la de 3 ori mai puține fiice bolnave decât de la tauri susceptibili la leucemie (Tabelul 65).
65. Incidența leucemiei la fiicele de tauri rezistente și susceptibile la leucemie (conform lui V.L. Petukhov)
Coeficientul de corelare a rangului taurinilor utilizați în diferite ferme a variat de la 0,40 la 0,75. Din tații a căror incidență a fiicelor a fost în intervalul de la 0 la 10%, s-au obținut fii care au dat naștere la urmași și cu o incidență relativ scăzută (5,4%). Cu o incidență mare a taților (30% sau mai mult), fiicele de fii au fost, de asemenea, caracterizate printr-o susceptibilitate crescută la leucemie (27,5). În ferme pot exista 21-28% dintre tauri cu o incidență relativ scăzută a bolilor (până la 5-10%) și 13-23% dintre tauri cu o incidență mare a descendenților (mai mult de 30%).
În cadrul fermelor individuale, au fost identificate diferențe între liniile de incidență a leucemiei. Cu toate acestea, rangul liniilor în diferite ferme și regiuni variază foarte mult. Doar 0,8-2,6% din varianța variabilității totale a trăsăturii se datorează influenței liniei. Au fost crescute linii de șoareci care diferă puternic în ceea ce privește incidența leucemiei și a altor neoplasme maligne.
Influența familiilor și a mamelor. Există diferențe mari între familii în ceea ce privește incidența leucemiei. Există familii care nu suferă de leucemie și cu o incidență de până la 70% (Ernst et al., 1989). În unele populații, au fost identificate până la 30% dintre familiile indemne de leucemie și până la 13% dintre familiile cu o incidență de peste 40%. Figura 66 prezintă o diagramă genealogică a unei familii susceptibile la leucemie.
Mamele cu leucemie au produs de 2 ori mai multe fiice bolnave decât fiice sănătoase (Tabelul 66). Incidența îmbolnăvirilor la nepoate depinde într-o măsură mai mică de starea de sănătate a bunicilor. Coeficientul de corelație pentru incidența leucemiei la mame și fiice este 0,2. Există o relație pozitivă între vârsta de debut a leucemiei la mame și fiice (r = 0,39). Unii tați variază și în funcție de vârsta la care apare leucemia la fiicele lor. Incidența leucemiei până la vârsta de 3 ani este mai mică decât incidența la vârste mai înaintate.
66. Incidența leucemiei la descendenți în funcție de starea de sănătate a părinților (conform lui V.L. Petukhov)
Concordanța gemenilor de același sex pentru leucemie este de 74%. Rudele apropiate ale gemenilor leucemici se îmbolnăvesc mai des decât cele mai îndepărtate.
A. G. Nezavitin (1992) în turme de reproducere au arătat că mamele RID (-) au primit de 2 ori mai puține fiice RID (+) decât mamele infectate cu VLCRS (Tabelul 67).
vnk ^n J£S?i2!Z?*** scheme * „Rechisiolomeinogo to leucemie semeispa Azartnaya L7M (primele cifre, nmshsshaye prin -nipe, indică anul rovdesha, ggorye -anul plecării, shshrvmer 72-75)
67. Frecvența infecției cu VLV a fiicelor obținute de la mame infectate și sănătoase (după A. G. Nezavitin)
Legătura cu sistemele polimorfe genetice. S-au dovedit diferite asocieri ale complexului major de histocompatibilitate cu anumite boli la oameni si animale. La bovinele Jersey, antigenele MHC W1 și W3 s-au dovedit a fi asociați cu rezistența la leucemie. Pe măsură ce animalele se infectează cu virusul leucemiei, frecvența antigenelor BoLA W8 și W11 crește în grupul de bovine hematologic-pozitive.
La vacile leucemice, alelele grupelor sanguine BG2O3Y2, Z sunt mai frecvente, iar la vacile sănătoase - BG2O3, BO3PFS2. Se crede că susceptibilitatea la leucemie este asociată cu anumite antigene ale sistemelor genetice ale grupelor de sânge.
Animalele brune letone cu ceruloplasmină de tip BB au o incidență mai mică a leucemiei (1,27%) decât animalele cu alte tipuri (Karlikov, 1976). Analizând toate datele disponibile privind asocierea sistemelor polimorfe cu leucemia, putem spune că nu există markeri genetici de rezistență și susceptibilitate a animalelor la leucemie.
Citogenetica leucemiei. Multe studii au descoperit modificări cariotipului în leucemie. În toate formele de leucemie bovină, este adesea detectat un procent crescut de aneuploidie. Un studiu al cariotipului măduvei osoase la animalele leucemice și sănătoase a arătat că bovinele bolnave au de 4 ori mai multe celule poliploide. Cu toate acestea, în unele studii nu există diferențe în frecvența anomaliilor cromozomiale la animalele bolnave și sănătoase. Nu a fost încă dovedit clar dacă anomaliile cromozomiale sunt una dintre cauzele leucemiei.
Moștenirea rezistenței și susceptibilității la leucemie. S-a demonstrat că moștenirea monogenă a rezistenței și susceptibilității la leucemie este puțin probabilă. Distribuția familiilor și a taurilor în ceea ce privește incidența leucemiei la descendenți a fost diferită de cea normală, în timp ce în aceleași efective incidența tuberculozei și brucelozei a fost normală. Unele studii au stabilit o distribuție normală a taurilor în ceea ce privește incidența leucemiei la fiicele lor. Se poate presupune că rezistența la leucemie este determinată de mulți loci genici. Coeficienții de heritabilitate de rezistență și susceptibilitate la leucemie în diferite efective variază de la 0,07 la 0,5 (Ernst, Petukhov, Karlikov etc.).
Se presupune că la pui, susceptibilitatea la infecția cu virusul leucemiei domină asupra rezistenței și este o trăsătură monogenă. Locii care determină rezistența împotriva virusurilor leucemice din subgrupele A și B sunt moștenite independent unul de celălalt.
Ipoteze și teorii ale etiologiei leucemiei. Cunoștințele despre neoplasmele maligne sunt în continuă expansiune. L.A. Zilber a creat o teorie virogenetică a apariției tumorilor, conform căreia acidul nucleic al virusului este inclus parțial sau complet. genomului celular.
O. A. Ivanova (1972) credea că leucemia este cauzată de un provirus (V), al cărui ADN este inclus în genomul unui animal disfuncțional. Genomul conține, de asemenea, o genă represoare (R), care afectează activitatea provirusului. Persoanele care au un provirus și o alelă recesivă inactivă a represorului Wrr, Vvrr și parțial WRr se îmbolnăvesc.
Leucemogeneza este un proces în mai multe etape. Se crede că leucemia este cauzată de virusul leucemiei bovine care conține ARN (BLV sau WmV), care aparține grupului de retrovirusuri. Acest virus este integrat cu genomul celulei hematopoietice și este capabil de transmitere orizontală și verticală.
În retrovirusuri, o oncogenă este responsabilă de inducerea cancerului. Oncogenele retrovirale nu sunt de origine virală, ci de origine celulară. Se presupune că o oncogenă (proto-oncogene) este o genă normală alterată. Când o proto-oncogenă intră în genomul unui retrovirus, aceasta este activată și transformată într-o oncogenă care poate transforma celulele. Există 17 proto-oncogene cunoscute. Cu toate acestea, nu au fost găsite oncogene în grupul de retrovirusuri ale leucemiei limfoide bovine.
V.P. Shishkov (1976) a prezentat un concept viral-imunogenetic al etiologiei patogenezei și prevenirii leucemiei și a unui număr de boli tumorale. Conform acestei teorii, alături de rolul principal al virusurilor care conțin ARN în dezvoltarea leucemiei la bovine și păsări, sunt importante starea imunologică a organismului și predispoziția genetică la această boală.
Reacția de imunodifuzie (IDR) este utilizată pe scară largă pentru a diagnostica leucemia. Multe efective pot avea un procent mare de animale infectate. Animalele infectate cu VLCV nu se îmbolnăvesc întotdeauna. Aceste animale mențin un nivel normal de productivitate. Cu toate acestea, 90-100% dintre animalele cu leucemie sunt infectate cu VLCRS. Selecția animalelor pentru rezistența la leucemie poate fi efectuată numai în efectivele cu niveluri ridicate de infectare a animalelor cu virusul leucemiei.
Cancer la ochi. Această boală a vitelor este comună în Australia. Se crede că razele ultraviolete sunt principalul factor de apariție a bolii. Rasele cele mai susceptibile la cancer de ochi sunt Herefords, Herefords sondați și încrucișările lor. Boala apare la animale la vârsta de 8-10 ani și foarte rar - până la 5 ani. Cancerul ocular nu a fost înregistrat la animalele cu pigmentare completă a pielii (Aberdeen Angus, Guernsey, Jersey, Swiss, etc.). La animalele cu pigmentare scăzută a irisului, boala apare mai des (Tabelul 68).
68. Cancerul ocular și pigmentarea irisului (conform lui Nishimura și colab.)
Dintre cele 353 de încrucișări Hereford x Charolais, 13,1% dintre animale nu au avut deloc pigmentare și, prin urmare, au avut o incidență mai mare a cancerului ocular.
Boala aftoasă. O boală acută virală extrem de contagioasă a artiodactililor. Zebui sunt relativ rezistenți la boală. Într-o fermă, din 1074 capete de vite, 97,1% s-au îmbolnăvit de febră aftoasă. Rata de incidență a animalelor din rasele Simmental și de stepă roșie este de 100%, maro caucazian este de 87,5, iar zebu-ul este de doar 20%. Există dovezi ale rezistenței familiale a animalelor la boală.
Boala Marek (MD) este o boală infecțioasă a păsărilor (cauzată de un virus care conține ADN), caracterizată prin contagiozitate, proliferarea țesutului limforreticular în organele interne, piele, mușchi și afectarea trunchiurilor nervoase periferice.
Unele rase de pui variază în ceea ce privește rezistența la boala Marek. Au fost de asemenea relevate diferențe interliniare (Tabelul 69). La multe rase au fost create linii rezistente de pui. Astfel, oamenii de știință englezi au creat o linie de pui Rhode Island cu rezistență 100% la boală. Liniile rezistente și susceptibile diferă și în ceea ce privește producția de interferon, care funcționează functie de protectie pentru infecții virale.
69. Rezistența la boala Marek a liniilor consangvinizate de pui și încrucișările lor după infecția experimentală (conform lui Gavora, Spencer)
Potrivit unor oameni de știință, rezistența la boli domină susceptibilitatea. În acest caz, rolul principal este jucat de 2-4 gene autozomale a trei loci. S-a stabilit o dominanță incompletă a rezistenței asupra susceptibilității. Genele legate de sex nu influențează semnificativ rezistența, deși mortalitatea cauzată de boală este mai mare în rândul femeilor decât în rândul bărbaților.
Există o relație destul de clară între alela B a grupului sanguin B 21 și rezistența la boala Marek (Tabelul 70). Aproape toți indivizii din linia N rezistentă au fost homozigoți pentru antigenul B21, în timp ce majoritatea indivizilor din linia P susceptibilă au avut antigenul B19.
70. Dependența rezistenței găinilor la boala Mvrek de antigenul B 21
Denumire: B~ - alte alele B, dar nu B 21.
Există un punct de vedere despre prezența a cel puțin două forme de rezistență genetică. Una dintre ele este asociată cu anumite alele ale sistemului de grup sanguin B, în special B 21, și este exprimată prin reactivitate imunogenetică crescută și rezistență legată de vârstă. A doua formă de rezistență este asociată cu loci Th-1 și Ly-4 și este determinată de rezistența celulelor limfoide împotriva infecției.
Multe anomalii cromozomiale au fost descrise la celulele infectate cu virusuri. În MD, aneuploidia și rupturile cromozomilor sunt anomaliile cel mai frecvent observate. Astfel, proporția metafazelor aneugoide în limfocite a fost de 10%, iar în celulele măduvei osoase - 12%, față de 1,7% la martor.
Unii cercetători cred că nu există o relație semnificativă între trăsăturile de performanță și rezistența la MD. Alte studii au găsit o corelație negativă între rezistența la MD și greutatea corporală a păsărilor de curte și greutatea ouălor, dar o corelație pozitivă cu producția de ouă.
Boala de Newcastle (pseudoplaga păsărilor) este o boală virală foarte contagioasă, în principal a găinilor. Se caracterizează prin pneumonie, encefalită și leziuni hemoragice în puncte multiple organele interne. În cazurile acute, mortalitatea în rândul animalelor tinere ajunge la 100%. Se găsește la găini diferențe genetice asupra rezistenţei şi sensibilităţii. În experimentele lui Frankis (1955), după infectare, rata mortalității puilor din familia A a fost de 30%, iar în familia D - 78%. În plus, mortalitatea după infecție a crescut mai rapid la indivizii din familia D.
Când puii au fost imunizați cu un vaccin care conținea un virus inactivat care provoacă boala Newcastle, s-au evidențiat diferențe foarte semnificative între familiile paterne. Coeficientul de ereditabilitate pentru puterea răspunsului imun a fost de 0,41.
Păsările lipsite de celule T sunt foarte susceptibile la boală. Cu agammaglobulinemie, puii sunt, de asemenea, sensibili la boală.
Prepelițele japoneze au fost folosite ca obiect model de selecție pentru rezistența la boala Newcastle. Ca urmare, incidența prepeliței în generațiile V-VI a scăzut de la 41,5 la 2,05 și 2,63%. Coeficientul de heritabilitate realizat este de 0,6. În grupul selectat pentru rezistență, mortalitatea a scăzut de mai multe ori, eclozionabilitatea și greutatea în viu a păsărilor au crescut.
Scrapie (scrapie) este o boală infecțioasă care se dezvoltă lent, care duce la modificări distrofice ale sistemului nervos central. Cauzat de un agent asemănător virusului, a cărui natură nu este clară.
Antigenele nu au fost încă găsite împotriva agenților patogeni ai infecțiilor lente (scrapie, visna, kuru etc.). Sunt afectate oile de 2,5-4,5 ani. Un semn caracteristic al bolii este mâncărimea. Oile își freacă lâna pe gard, lăsând pete goale pe trup. Boala apare pe toate continentele și în multe țări, în special Australia, Noua Zeelandă, Africa de Sud și SUA. Europa de Vest scrapie a fost înregistrată de 200 de ani.
Oile din rasele Ramboulier, Hampshire și Targhee sunt mai rezistente la scrapie (14, 18 și, respectiv, 19%) decât oile Suffolk din Noua Zeelandă (39%) și punctele negre scoțiene (28-43%). Caprele de lapte sunt mai predispuse la scrapie (61%) decât caprele Angora (26%).
Selecția liniilor de rase de oi englezești Cheviot și Herdwick a fost eficientă pentru creșterea (linia pozitivă) și scăderea (linia negativă) a incidenței scrapiei ca răspuns la administrarea subcutanată a agentului scrapie, denumit SSBP// (Tabelul 71).
71. Scrapie la oi cauzată de infecția cu SSBP // (citat de R. Kimberlin)
Perioada de incubație la animalele de pe linia negativă este de multe ori mai lungă decât la animalele de pe linia pozitivă. Se crede că răspunsul la infecție este controlat de o genă numită sip (scrapie, perioada de incubație) care are două alele. Alela dominantă controlează susceptibilitatea la scrapie, în timp ce alela recesivă controlează rezistența. Trebuie luat în considerare faptul că răspunsul unei filiații la diferiți agenți scrapie nu este același. De exemplu, incidența oilor Cheviot a liniilor pozitive și negative pentru agentul scrapie SSBP// a fost de 100 și, respectiv, 50%. Incidența acestor linii în timpul infecției intradermice cu agentul scrapie CH 1641 a fost diferită: 53, respectiv 38%. Cu toate acestea, rata de morbiditate a animalelor de linie negativă în ambele cazuri a fost mai mică decât cea a animalelor de linie pozitivă. Acest lucru sugerează posibilitatea de a crea linii de animale rezistente la mai multe tulpini ale unui agent patogen. Controlul genetic al rezistenței la scrapie depinde de octgama agentului patogen.
Mixomatoza de iepure. O boală virală acută caracterizată prin conjunctivită sero-purulentă și formarea de tumori la nivelul capului, anusului și organelor genitale externe. La unele rase de iepuri, ratele mortalității ajung la 100%. Posibilitatea creșterii proporției de rezistență a indivizilor dintr-o populație este evidentă din exemplul următor. În 1859, 3 iepuri importați din Europa au fost eliberați în Australia. În 1953, erau aproximativ 0,5-1 miliarde de ei agricultură. În 1950-1951 în unele zone din Australia, iepurii au fost infectați cu virusul mixomatozei, iar după un timp până la 90% dintre animale au murit. Cu toate acestea, problema nu a fost rezolvată, deoarece printre iepuri au apărut indivizi rezistenți genetic, iar numărul acestora a crescut foarte mult. Selecția pentru rezistența la mixomatoză la rasele de iepuri domestice a avut, de asemenea, succes. Coeficientul de heritabilitate al rezistenței la această boală este de 0,35-0,40.
Biletul 47.
Se știe că aproape toate reacțiile organismului (normale și patologice) sunt determinate de fondul genotipic individual. Factorii genetici, având semnificaţie etiologică, determină desfăşurarea procesului patologic numai atunci când acţionează unidirecţional împreună cu un anumit set de factori exogeni (de mediu extern).
Boala ulcerului peptic este, în general, o boală multifactorială cu un tip de moștenire poligenică - este o boală cu predispoziție ereditară. Din punct de vedere genetic, boala ulceroasă peptică este un grup eterogen de boli care diferă prin gradul de încărcare genetică, cea mai gravă formă a bolii fiind forma copilăriei a bolii, iar la adulți, localizarea ulcerului peptic în zona piloroduodenală este cea mai gravă. Descoperirea sindroamelor de ulcer peptic monogenic indică faptul că conceptul de „boală ulceroasă peptică heterogenă genetică” include nu numai forme ale bolii cu diferite grade de povară genetică, ci și forme cu diferite tipuri moștenire, deși trebuie presupus că numărul de astfel de forme monogenice de ulcer peptic este probabil nesemnificativ, iar în cea mai mare parte a cazurilor de ulcer peptic sunt de natură polietiologică, adică. sunt realizate prin acţiunea combinată a factorilor genetici şi de mediu.
Componenta ereditară în boala ulcerului peptic (în special în cazul localizării piloroduodenale a ulcerului) este probabil asociată cu un număr semnificativ de gene, fiecare dintre acestea determinând în sine prezența unei trăsături „normale” și în combinație cu alte gene similare. trăsăturile contribuie la atingerea unui anumit nivel „prag” - prezența susceptibilității genetice la boala ulcerului peptic.
Asociațiile de încredere identificate ale bolii cu caracteristicile mendeliane fac necesar să se considere aceste caracteristici ca factori de risc pentru dezvoltarea bolii ulcerului peptic. Factorii genetici dovediți includ indicatori ai secreției maxime de acid clorhidric; conținutul de pepsinogen I în serul sanguin; eliberare crescută de gastrină ca răspuns la aportul alimentar.
Factorii despre care se crede că au o bază ereditară în dezvoltarea bolii includ următorii:
2. eliberare crescută de acid clorhidric după masă;
3. sensibilitate crescută a celulelor parietale la gastrină;
4. încălcarea mecanismului feedbackîntre producția de acid clorhidric și eliberarea de gastrină;
5. tulburări ale funcției motorii ale stomacului și duodenului;
6. activitate scăzută a enzimei Z1-antitripsină;
8. încălcarea producției de imunoglobuline A;
9. modificări morfologice ale mucoasei (gastrită, duodenită) și altele.
Markerii genetici pentru dezvoltarea ulcerului gastric și duodenal sunt anumite grupe sanguine și caracteristici fenotipice.
Un loc semnificativ printre factorii genetici ai bolii ulcerului peptic este acordat capacității de a secreta antigene de către sistemul AVN, detectarea antigenului HZA și histocompatibilitatea B5, B15, B35.
În prezent, pe baza factorilor ereditari și constituționali, a fost formulată o teorie eterogenă, conform căreia ulcerogeneza se bazează pe polimorfismul ulcerului peptic, o combinație de diverși factori predeterminați genetic care determină polimorfismul manifestărilor clinice ale bolii ulceroase peptice. .
Biletul 48.
În creșterea animalelor industriale, bolile membrelor sunt cele mai frecvente boli. De la 3 la 7% din pierderea de lapte este asociată cu bolile copitei la vaci. În unele țări, sacrificarea din cauza bolilor membrelor este de 2,4-15,0%. La bovinele olandeze, 6-7% dintre vacile de lapte sunt afectate de defecte ale copitei care provoacă șchiopătare. Animalele se îmbolnăvesc cel mai adesea la vârsta de 4-7 ani.
Zeci de boli ale membrelor sunt adesea grupate sub denumirea generală „boli ale membrelor”. Condițiile de mediu joacă un rol major în apariția lor, dar ereditatea este adesea un factor predispozant important. Au fost descrise o serie de boli ereditare monogenice ale membrelor, dar bolile cu predispoziție ereditară ocupă un loc foarte mare.
Rasele gri locale și bulgare de bovine se remarcă prin constituția lor puternică și în special prin picioare și copite puternice. Incidența vacilor din rasa Estonă alb-negru este de 2 ori mai mare decât cea a rasei Estoniene roșii. A fost studiată rezistența copitelor vitelor alb-negru și încrucișările sale cu Holstein. Încrucișările de tauri suedezi au fost caracterizate de cea mai mare rezistență a copitelor. Cea mai scăzută rezistență a copitei a fost găsită în încrucișările de tauri Holstein din SUA, Noua Zeelandă și Olanda.
Vitele alb-negru din Siberia sunt mai puțin susceptibile la complicațiile infecțioase ale membrelor (11,3%) decât
bovine importate daneze și olandeze (21, 30,4%) și încrucișările acestora (Tabelul 74).
Au fost relevate diferențe semnificative în incidența morbidității la membrele fiicelor de tauri alb-negru. Astfel, într-unul dintre efective, incidența urmașilor taților individuali a variat de la 0 la 23%, cu o incidență medie de 9% pentru efectiv. Coeficientul de ereditabilitate pentru forța generală a membrelor este 0,13. Vitele din rasele alb-negru (tip lapte-carne) și Hereford sunt mai puțin predispuse la boală.
Diversitatea genetică în rezistența la boli ale extremităților de diferite forme nu este aceeași. De exemplu, h2 de paralizie a membrelor posterioare este 0,3, iar tumorile despicaturii intercopete (limax) este 0,2.
Frecvența limaxului la fiicele diferiților tauri a fost în intervalul 11-53%, cu o medie a efectivului de 23%. Diferențele dintre linii sunt nesigure. Nu s-au găsit diferențe semnificative în frecvența deformărilor și a complicațiilor inflamatorii și infecțioase între animalele consangvinizate și cele extrasange La vacile cu antigenul grupului sanguin J s-au găsit mai puține complicații inflamatorii și infecțioase (4,4%) decât la animalele cu antigenul Oo (19,5). 96).
Ereditatea joacă un anumit rol în rata de creștere, rezistența la abraziune, numărul de tuburi de corn pe unitatea de suprafață a copitelor, capacitatea de absorbție a apei a cornului copitei etc. Astfel, heritabilitatea numărului de tuburi de corn este de 0,38-0,75, diametrul tuburilor este de 0,63, gradul de abraziune a copitei este de 0,79 și capacitatea de umiditate este de 0,82. Leziunile copitei sunt asociate cu o predispoziție ereditară la formarea cornului elicoidal și a ulcerului tălpii.
Amplasarea membrelor poate influența, de asemenea, morbiditatea. De exemplu, cu o poziție în formă de sabie, se observă ulcere și supurație, formarea de copite în unghi acut sau plate cu o poziție în formă de X, poate apărea laminită purulentă abaxială.
Pareze spastică (sindrom) (lezarea membrelor pelvine). Boala se caracterizează prin moștenire poligenică recesivă cu penetrare incompletă a factorului recesiv. În cele mai multe cazuri, factorul recesiv se transmite prin tauri. Sindromul spastic poate apărea la 10-30 de tauri la stațiile de inseminare artificială
Incidența bolii este deosebit de mare la taurii Holstein și Aberdeen Angus. Datorită utilizării pe scară largă a taurilor Holstein în țara noastră, este necesar să se țină cont de forme precoce (3,5-6 luni) și tardive (3-8 ani) de sindrom spastic și alte boli în munca de reproducere. În SUA, 2,6% din cei 12.387 de tauri au avut cinci produse lactate și trei rase de carne au fost respinse din cauza sindromului spastic. Mai mult, incidența bolii a fost mai mare la taurii din rasele Ayrshire și Holstein (3,9, respectiv 3,5%). În unele țări, 5-10% dintre tauri au fost sacrificați din cauza acestei boli.
Există opinia că utilizarea consangvinizării crește incidența la 12-17%.
La trei generații de tauri ai vitelor pestrițe maghiare din linia Sultana, forma clinică de pareză spastică a fost înregistrată la 7 din 10 tauri. 1028 de descendenți ai taurilor călcâi din linia Sultan au fost studiați în trei generații. Rata de incidență a puilor de tauri bolnavi clinic în Fj este de 43,5%, în F2 - 31,7 și F3 - 31,3%. Incidența bolii la descendenții de tauri clinic sănătoși a fost de câteva ori mai mică: în Fi - 29,1%, în F2 - 14 și în F3 - 6,6%. Incidența parezei spastice în unele familii a ajuns la 60%. Când sunt ținute în boxe, pareza spastică de formă tardivă (3-8 ani) a fost observată la 58% dintre animale, iar când au fost ținute pe pășune - la 42% (Hamori, 1983).
Au fost studiate 13 semne de slăbiciune a picioarelor la porci. Coeficienții de ereditabilitate pentru aceste trăsături au variat de la 0 la 0,56, iar h2 medie pentru slăbiciunea picioarelor a fost de 0,07. Studiile cu raze X ale porcilor bacon au arătat că h2 de osteocondrită a membrelor este de 0,4. Există diferențe între pui și între descendenții diferiților mistreți în incidența slăbiciunii picioarelor și a leziunilor articulațiilor.
Diferite rase de oi variază în ceea ce privește susceptibilitatea la putregaiul piciorului cauzat de Fusiformis nodosus. Rata de incidență a Coimbatores de rasă pură în India a fost de 12,8%, iar cea a Corridelilor a fost de 61,1%. La hibrizii Fi și 3/4 sânge conform coridelilor, putregaiul piciorului a fost înregistrat la 21,1 și, respectiv, 26,1% dintre oi. În toate loturile, incidența bolilor la oile adulte a fost de câteva ori mai mare decât la miei. Membrele anterioare sunt mai des afectate decât membrele posterioare. Se poate presupune că rezistența la putregaiul picioarelor nu domină complet susceptibilitatea. Rasele britanice de oi și încrucișările lor sunt mai rezistente la boală decât oile Merino.
Un studiu al prevalenței piciorului roșu congenital la porcii Landrace, Yorkshire și Duroc a arătat că la vierii Landrace incidența piciorului roșu congenital a fost mai mare decât la animalele din alte rase. Frecvența piciorului roșu congenital în rândul descendenților diferiților mistreți a variat de la 0 la 22%. Mai mult decât atât, incidența piciorului roșu la mistreți este de 1,7 ori mai mare decât la scrofe. Greutatea corporală a purceilor cu picior roșu la naștere a fost mai mică decât a purceilor sănătoși. S-a făcut o concluzie despre cauza poligenică a piciorului roșu congenital.
Organizarea diagnosticului diferențiat al bolilor membrelor, evaluarea descendenților tarilor și familiilor pentru rezistența la aceste boli este un element necesar al muncii de reproducere.
Biletul 49.
Infertilitatea este o încălcare a reproducerii descendenților. Infertilitatea completă sau parțială a fost raportată la 65% dintre tauri din Suedia. În SUA și Canada, din 8887 de tauri de lapte, 36% au fost sacrificați din cauza diverse încălcări capacitatea de reproducere. La 20-40% dintre vaci apar diferite forme de infertilitate. La 32 de efective de vaci Holstein din SUA, metrita a fost înregistrată la 18,2% dintre animale, chisturi ovariene în 10,4, placenta reținută în 8,6, avorturi la 1,4% dintre animale. La bovinele roșii norvegiene, corelațiile fenotipice ale placentei reținute, metritei, febrei laptelui, mastitei, cetozei cu infertilitate sunt respectiv 0,77; 0,73; 0,69; 0,68; 0,57. Aceasta înseamnă că toate aceste boli afectează semnificativ apariția infertilității.
Gradul de manifestare a capacității de reproducere variază de la fertilitate normală la infertilitate absolută. Încălcarea capacității de reproducere este cauzată de numeroși factori de mediu și de ereditate. Cu toate acestea, este imposibil să distingem clar între tulburările de fertilitate neereditare și cele ereditare.
Coeficienții de ereditabilitate ai infertilității sunt foarte mici - de la 0 la 0,1 (Zavertyaev, 1986). Cu toate acestea, diferențele în incidența infertilității la descendenții diferiților tauri variază de la 0 la 30%.
Calitățile de reproducere ale vacilor Holstein sunt oarecum mai rele decât cele ale vacilor albe și negre. Vacile Holstein necesită 1,91 de inseminare pe fătare, vacile albe și negre necesită 1,55 iar vacile roșii și albe necesită 1,59.
Infertilitatea este frecventă la junincile albe (boala junincilor albe) de la Shorthorn și la rasele înrudite. La junincile albe, uterul si vaginul sunt subdezvoltate, in timp ce ovarele si organele genitale externe sunt dezvoltate normal. Această formă de infertilitate apare la 10% dintre junincile albe Shorthorn. Într-un efectiv de Shorthorn, incidența infertilității la junincile roșii, roan și albe, respectiv, a fost de 1,1; 3,5 și 39,1%.
Vitele albe și albastre din Belgia Mijloc Superioară provin din Shorthorns. Procentul de juninci infertile la bovinele albe (14,9) este de 3 ori mai mare decât la bovinele albastre (4,2).
De asemenea, s-a stabilit că printre junincile albe, 10-15% sunt infertile, dintre junincile roan - 1-1,75, iar dintre junincile negre - doar 0,3%.
Hipoplazia (subdezvoltarea) a ovarelor și a testiculelor a fost observată anterior adesea la bovinele strânse din nordul Suediei (aproximativ 17%). Juincile cu hipoplazie bilaterală nu intră în căldură, iar taurii cu hipoplazie gonadală bilaterală nu produc spermatozoizi.
La bovinele suedeze Fjäll, a fost găsită o relație între frecvența hipoplaziei și prevalența colorației albe pe suprafața corpului. La animalele cu o prevalență a colorației albe de 0,9 părți ale suprafeței corpului, incidența hipoplaziei gonadale a fost de 13,5%, iar la animalele cu pigmentare de peste 50% din suprafața corpului nu a existat nicio boală. În consecință, un semn calitativ, într-o oarecare măsură, poate fi un marker al unor forme de infertilitate.
Printre defectele organelor sistemului reproducător, trebuie menționat criptorhidia, în care unul sau două testicule se află în cavitatea abdominală și hernia scrotală, în care, datorită diametrului mare al inelului inghinal care leagă cavitatea abdominală cu scrot. , ansele intestinale se extind în scrot. Criptorhidia este mai puțin frecventă la bovine decât la cai, capre, porci și oi. La armăsarii cu criptorhidie unilaterală rata de fertilitate a iepelor a fost de 23%, iar la cei normali a fost de 53%. Criptorhidia este cea mai frecventă la câinii din rase cu craniul scurtat (până la 23% la boxeri). La oile de rasă prekos cu lână fină, proporția de criptorhide (în mare parte bilaterale) între berbeci ajunge la 25%. La porci, criptorhidia apare la 0,2-0,4% dintre indivizi, iar la unele populații - până la 3-5%.
Criptorhidia se poate răspândi rapid în populațiile de oi. La unele ferme din Ungaria, incidența criptorhideei a ajuns la 25-30%. În rândul berbecilor cu corn, criptorhidia a fost înregistrată la 75,6% dintre indivizi, iar la berbecii cu coarne - 2,4%. Hipoplazia testiculelor a fost observată la 9 berbeci din 141. În rândul animalelor urcate, hipoplazia a fost la 77,7%, la animalele cu coarne - la 11,5%. La capre și porci, criptorhidia este moștenită în mod autosomal recesiv. Studiile efectuate în SUA pe porci polonez-chinezi și Landrace au arătat că coeficientul de ereditabilitate al herniei scrotale este de 0,15. Ponderea influenței materne este aproximativ egală cu aceeași valoare. Criptorhidia și hernia scrotală sunt caracteristici limitate de gen.
Hermafroditismul (combinația de sex masculin și feminin într-un singur organism) este mai frecvent la capre și porci și mai puțin frecvent la bovine, cai și oi.
Toți hermafrodiții au cromozomi sexuali XX. Unii indivizi au doar țesut testicular, alții au țesut testicular și ovarian, dar nu se formează celule sexuale. Frecvența hermafrodiților variază între diferite linii și rase. La un porc alb cu urechi lungi este de 0,1%.
Intersexualitatea este mai frecventă la caprele cocotate. Într-una dintre efectivele de capre Saanen și Toggenburg au fost identificate 11 și, respectiv, 6% dintre indivizii intersexuali. Femelele erau genetic intersexuale. În Suedia, au fost identificați 0,21% dintre porcii intersexuați. În Statele Unite, un studiu a raportat că 0,53% erau intersexuali. La Landrace frecvența unor astfel de porci este de 1,5%, iar la rasa White Bacon din Ungaria este de 3,8%. Animalele intersexuale sunt supuse sacrificării.
Incidența nașterii mortii și a avortului în unele efective poate varia foarte mult (de la 1 la 10% și, respectiv, de la 0,5 la 5% sau mai mult) și se datorează multor factori genetici și non-genetici. Există fenomene imunitare la toate nivelurile de reproducere normală în corpurile femelelor și bărbaților. Unul dintre motivele pierderilor prenatale poate fi o stare de imunodeficiență congenitală sau dobândită a femeilor. Coincidența anumitor antigeni ai complexului major de histocompatibilitate poate provoca perturbarea dezvoltării embrionare. La porci, în 8,5% din cazuri, moartea embrionului se observă în primele 15-20 de zile după fecundare.
Există diferențe încrucișate în ceea ce privește incidența nașterilor morti, avorturilor și fătărilor dificile (Tabelele 75, 76). Vitele roșii daneze au un procent semnificativ mai mare de fătări dificile și născuți morti decât vitele Angler. Cu toate acestea, junincile primului vițel au o rată mai mare de fătări dificile și de nașteri morti decât vacile. Corelația genetică dintre fătări dificile și mortalitatea vițeilor a fost de 0,66. La junincile Holstein din Statele Unite, 1% dintre animale au reținut placenta și 45% au avut fătări dificile. La junincile prim-vițel din rasa Ayrshire, retenția de placentă a fost de multe ori mai mare (12-22%), iar numărul fătărilor dificile a fost mai mic (8-18%). Pentru a reduce frecvența fătărilor dificile, este necesar utilizați tauri cu risc scăzut de apariție a acestei tulburări.
75. Frecvența vițeilor născuți morți și a avorturilor la vaci (conform lui V.L. Petukhov)
76. Frecvența nașterilor morti și a fătărilor dificile la unele rase din țările scandinave și Germania (citat de Dekhtyarev și colab.)
S-a stabilit că frecvența avorturilor este mai mare la fiicele ale căror mame erau predispuse la avort. Coeficienții de ereditabilitate pentru predispoziția la avort și nașterea mortii au fost în intervalul 0-0,08, pentru fătarea dificilă - 0,04-0,30. S-a dovedit o influență semnificativă a genotipurilor taurului asupra frecvenței acestor trăsături.
Mortalitatea prenatală la ovine este de 5-23%. Rate mai mari de mortalitate apar la gemeni.
Una dintre metodele de combatere a infertilității este evaluarea genotipului producătorilor în funcție de acest indicator și utilizarea pe scară largă a celor mai bune dintre ele.
Concepte de bază.Rezistenţă- rezistența organismului la acțiunea agenților fizici, chimici și biologici care provoacă o afecțiune patologică. Acest concept este oarecum mai larg decât imunitatea, deși uneori sunt folosite ca sinonime.
Susceptibilitate- predispoziţia organismului la efectele fizice, chimice şi factori biologici conducând la o stare patologică.
Rezistența și susceptibilitatea la animalele din aceeași specie, de regulă, nu sunt absolute, ci relative. Poate fi mare, medie și scăzută.
Moștenirea rezistenței și susceptibilității. Natura genetică a bolilor cu predispoziție ereditară a fost puțin studiată. Acest grup de boli se caracterizează prin: 1) controlul polifactorial (determinat de mulți loci) al rezistenței și susceptibilității; 2) influența condițiilor de mediu; 3) tranziție continuă de la forme pronunțate ale bolii la normal, adică de la susceptibilitate la rezistență; 4) prevalență ridicată, diferențe genetice minore între populații; 5) variabilitate mare a vârstei de manifestare a bolii; 6) adesea puțină concordanță în perechi de gemeni identici.
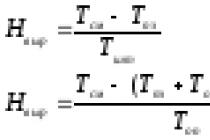
În funcție de fenotip, animalele cu privire la bolile cu predispoziție ereditară pot fi împărțite în două clase: sănătoase și bolnave. Animalele se îmbolnăvesc atunci când ating pragul adecvat T pentru acțiunea alelelor active și un anumit nivel de condiții de mediu. Rezistența sau susceptibilitatea se referă la caracteristicile pragului - Acestea sunt caracteristici a căror distribuție în timpul divizării are loc intermitent, dar sunt moștenite polifactorial. Trebuie amintit că animalele susceptibile ereditar nu se îmbolnăvesc dacă nu există un agent patogen virulent.
Metode de studiu a rezistenței ereditare și a susceptibilității la boli. Există mai multe abordări principale pentru studierea determinării genetice a rezistenței și susceptibilității animalelor la boli: 1) analiza clinică și genealogică. Pentru efectuarea analizei clinice și genealogice se întocmesc diagrame genealogice ale familiilor și liniilor, indicând toate cazurile de boli. Se calculează frecvența morbidității în cadrul grupurilor înrudite, prin care acestea sunt comparate între ele și cu frecvența populației.
; 2) analiza gemeni. Această metodă face posibilă determinarea rolului relativ al eredității și al mediului în etiologia bolii. În acest scop, se determină concordanța și discordanța. Concordanţă- prezența sau absența bolii la ambii gemeni și discordanţă- un fenomen în care doar un geamăn are acest semn.
3) identificarea diferențelor de rasă și interlinie; . Analiza acestor diferențe de rezistență la boli indică rolul factorilor genetici în determinarea acestei trăsături.
4) experiment de selecție; Dacă, ca urmare a selecției, rezistența la o boală crește, aceasta indică faptul că rezistența și susceptibilitatea sunt determinate genetic.
5) analiza statistică a populației; Este folosit pentru a studia genetica rezistenței și susceptibilității bolilor multifactoriale, precum și în studiul trăsăturilor utile din punct de vedere economic.
6) analiza legăturii dintre boli și genele marker etc. Analiza acestei conexiuni este o altă modalitate de a demonstra determinarea ereditară a rezistenței și susceptibilității la boli.
Când se studiază rezistența și susceptibilitatea ereditară, nu una, ci o combinație a acestor metode este utilizată în diferite combinații.
Moștenirea simplă a stabilității. Acest termen înseamnă că rezistența la o boală este controlată de una sau câteva gene. În prezent sunt cunoscute doar câteva boli ereditare-de mediu cu moștenire simplă.
V i r u s s. Moștenirea autosomal dominantă a rezistenței la leucemia limfoidă a fost descrisă la pui. În acest caz, se disting două niveluri de rezistență genetică: rezistența celulară la infecție viralăși rezistența la dezvoltarea tumorii la păsările infectate cu virus.
Bacteriile. Unele tulpini de Escherichia coli (E. coli) provoacă diaree (diaree) la porcii nou-născuți. Cu toate acestea, unii purcei pot fi rezistenți la tulpinile de E. coli care au antigene K-88. Această rezistență este cauzată de receptorii care recunosc antigenele K-88. Prezența receptorilor este o trăsătură ereditară simplă.
Nematode. Există dovezi că rezistența oilor la hemonhoză este moștenită ca o simplă trăsătură dominantă.
Rezistență genetică și susceptibilitate la boli bacteriene (mastita, bruceloză, tuberculoză, leptospiroză)
Mastită- inflamatia glandei mamare. Cauzele bolii pot fi factori biologici (streptococi), mecanici, termici si chimici. Diferențele de încrucișare Nu există doar incidența mastita, ci și rezistența la agenți patogeni individuali. Bivolii au o incidență mai mică a mastitei decât vacile de lapte. Influența producătorilor, liniilor și familiilor. Există diferențe mari de incidență între fiicele taților diferiți. La taurii rezistenți, 3-15% dintre fiice se îmbolnăvesc, iar la taurii susceptibili - 20-50% și mai mult. Influența producătorilor asupra rezistenței puilor la mastită este de 10--19. Forma ugerului și tetinelor. Vacile cu uger în formă de cupă și rotunjite și o dispoziție calmă sunt mai puțin susceptibile de a dezvolta mastită. Într-o oarecare măsură, susceptibilitatea la boală este influențată de uniformitatea dezvoltării sferturilor ugerului, de forma și dimensiunea mameloanelor. Animalele cu ugerul căzut sunt mai susceptibile la mastită. Productivitatea laptelui. Vacile cu o rată mare de producție de lapte sunt mai susceptibile la mastită decât vacile cu o rată medie de producție de lapte. Cu o creștere a producției zilnice de lapte cu 1 kg, tulburările în secreția de lapte cresc cu 2%. Odată cu vârsta, incidența mastita crește. În timpul celei de-a doua și a treia fătări, incidența bolii poate crește. Numărul celulei somatice se coreleaza cu incidenta mastita. Vacile cu un număr scăzut de celule de lapte în prima lor lactație au fost mai puțin susceptibile de a dezvolta mastită în lactațiile ulterioare. Bruceloză-- o boală infecțioasă cronică a animalelor și a oamenilor cauzată de bacterii. La multe animale se manifestă ca avort, placentă reținută și tulburări de fertilitate. Diferențele de specii și rasă. Cobai, șoarecii albi și gophers sunt cei mai sensibili, în timp ce șobolanii albi, gâștele și porumbeii sunt rezistenți. Influența producătorilor, liniilor și familiilor. Influența producătorilor asupra incidenței bolilor la descendenți este de 8%. Nu au fost stabilite diferențe între liniile de rezistență și susceptibilitate a animalelor la bruceloză. În familiile de gemeni de același sex, gemenii afectați proveneau în principal din familii în care incidența a fost de 44%, iar gemenii sănătoși proveneau din familii cu o incidență de 20%. Moștenirea rezistenței și susceptibilității la bruceloză. Pentru mulți boli infectioase caracterizat prin absenţa unor grupuri mari înrudite de animale cu stabilitate absolută. Rezistența la bruceloză este controlată de o genă autosomal dominantă, iar susceptibilitatea de o genă recesivă. Tuberculoză- boala infectioasa. Agentul cauzal este micobacteriile. O boală a mamiferelor, păsărilor și oamenilor, caracterizată prin formarea în diferite organe a tuberculilor tipici - tuberculi, care suferă necroză cazeoasă. Această boală provoacă daune enorme producției animale și reprezintă o amenințare pentru sănătatea umană. Diferențele interspecifice și de rasă. Crescătorii de animale și porcii sunt cei mai sensibili la tuberculoză. Caprele și câinii se îmbolnăvesc mai rar. Caii și pisicile sunt relativ rezistenți la infecții. Influența producătorilor de linie și familie. Coeficientul de corelație dintre incidența tuberculozei la fiicele taților și fiicele fiilor acestora este de 0,33. Influența producătorilor asupra incidenței tuberculozei la descendenți este de 6%, iar cea a familiilor este de 25%. Moștenirea rezistenței și susceptibilității. Un factor important care determină rezistența ereditară congenitală și dobândită la tuberculoză este capacitatea macrofagelor de a suprima creșterea bacteriilor în citoplasma lor. Leptospiroza- boală focală naturală infecțioasă a animalelor și a oamenilor. Agentul cauzal este Leptospira. La animale se manifestă ca febră, anemie și avort. La încrucișarea animalelor rezistente și susceptibile, s-a observat o dominanță incompletă a rezistenței la leptospiroză.
Mastita este o inflamație a glandei mamare. Cauzele bolii pot fi biologice (streptococi, stafilococi etc.), mecanici, termici si chimici. La vacile bolnave, producția de lapte scade în medie cu 300 kg pe an. În unele țări, incidența ajunge la 30-50%. Pierderile anuale cauzate de mastită în Statele Unite se ridică la peste 2 miliarde de dolari.
Diferențele de încrucișare există nu numai în ceea ce privește incidența mamitei, ci și în rezistența la agenții patogeni individuali. La bovinele Holstein incidența este de până la 58%, la bovinele Guernsey - 33,6, Ayrshire - 5,6, rasa Harian - 2%. Bivolii și zebui au o incidență mai mică a mastitei decât vacile de lapte. Animalele din rasa Iskar sunt mai rezistente (13%) decât vacile din rasele brune bulgare și Simmental (25-28%), iar la bivoli 0,5% dintre indivizi s-au îmbolnăvit. Vitele Ayrshire au prezentat, de asemenea, o incidență mai mică (4,1%) decât vacile Kholmogory (12,3%) și Brown Latvian (29%). În același timp, vacile Ayrshire sunt mai susceptibile la mastită decât vacile finlandeze.
Influența producătorilor, liniilor și familiilor. Există diferențe mari de incidență între fiicele diferiților tați (A.I. Zheltikov, 1992). La taurii rezistenți, 3-15% dintre fiice se îmbolnăvesc, iar la taurii susceptibili - 20-50% și mai mult. Influența producătorilor asupra rezistenței puilor la mastită este de 10-19%. Unii cercetători au găsit diferențe între liniile de rezistență și susceptibilitate la mastită, alții nu au stabilit astfel de diferențe. Coeficientul de heritabilitate de rezistență și susceptibilitate a vacilor la mastită este în intervalul 0,05-0,80. Coeficientul de repetabilitate al costum-t este 0,36.
S-au evidențiat diferențe mari în incidența mastitei la animale între familii (0-50%). În unele efective, mamele cu mastită produc de 1,5-2 ori mai multe fiice bolnave decât mamele sănătoase.
Forma ugerului și tetinelor. Vacile cu uger în formă de cupă și rotunjite și o dispoziție calmă sunt mai puțin susceptibile de a dezvolta mastită. Într-o oarecare măsură, susceptibilitatea la boală este influențată de uniformitatea dezvoltării sferturilor ugerului (incidența sferturilor posterioare este mai mare), de forma și dimensiunea mamelonelor. Animalele cu ugerul căzut sunt mai susceptibile la mastită. Vacile cu tetine cilindrice sunt mai puțin afectate decât cele cu tetine în formă de pâlnie.
Portiunea distala (4-5 mm) a canalului mamelon este prima bariera de protectie impotriva agentilor patogeni datorita factorilor mecanici si a unor substante antimicrobiene. Proprietățile antibacteriene ale epiteliului canalului mamelonului sunt determinate de concentrația mare de acizi grași din celulele epiteliului care conțin cheratină - lauric, miristic și palmitic.
Nivelurile ridicate de acizi grași stearic, linolenic și oleic cresc susceptibilitatea la mastită. Se crede că heritabilitatea ridicată a acizilor grași (0,38-0,58) face posibilă utilizarea acestora ca criteriu de selecție pentru rezistența la mastită. În țesuturile interne de la intrarea în canalul mamelonului a fost detectată o concentrație de celule plasmatice care participă la protejarea ugerului de infecție.
Polimastia (sfarcurile suplimentare) este, de asemenea, un factor predispozitiv la mastita. Coeficientul de ereditabilitate al polimastiei este de 0,56.
Productivitatea laptelui. Vacile cu o rată mare de producție de lapte sunt mai susceptibile la mastită decât vacile cu o rată medie, dar există dovezi că producția lentă de lapte este un factor predispozant. Există o corelație pozitivă între incidența mamitei și producția de lapte (r = 0,20 - 0,30). Odată cu o creștere a producției zilnice de lapte cu 1 kg, tulburările de secreție de lapte cresc cu 1,8%. Cu toate acestea, mulți cercetători nu au stabilit o relație între nivelurile de producție de lapte și mastita. În același timp, la mulți tauri nu există o legătură sigură între mastită și producția de lapte a fiicelor. Vacile cu viață lungă și foarte productive sunt mai puțin probabil să fie sacrificate din cauza mastitei decât vacile cu viață scurtă. Odată cu vârsta, incidența mastita crește. La a doua și a treia fătare, incidența bolii poate crește de 2-3 ori față de prima.
Factorii care protejează glanda mamară de infecții sunt leucocitele polimorfonucleare, sistemul lactoperoxidază, lizozima, lacgoferina (Lf), sistemul complementului.
Există diferențe semnificative între vaci în ceea ce privește capacitatea neutrofilelor polimorfonucleare din lapte de a fagocita stafilococi. Variabilitatea genetică a activității fagocitare a fost dovedită în experimente pe șoareci. Activitatea fagocitară se corelează cu rezistența vacilor la mastită (r = 0,40).
Numărul de celule somatice se corelează cu incidența mastitei (r = 0,28). Vacile cu un număr scăzut de celule de lapte în prima lor lactație au fost mai puțin susceptibile de a dezvolta mastită în lactațiile ulterioare. Cu un număr de celule de până la 500 mii/ml, incidența a fost de 30-38%, iar cu un număr de celule peste 500 mii/ml - 56%.
Lizocimul laptelui are efect bacteriostatic și bactericid. La concentrații mari în lapte are loc o scădere mai rapidă a numărului de bacterii patogene decât la concentrații scăzute. Concentrația de lizozim în lapte în timpul mastitei crește de 3-7 ori. Au fost dezvăluite diferențe de încrucișare și intrabreed între animale în conținutul de lizozim.
Activitatea bacteriostatică este, de asemenea, caracteristică lactoferinei. Severitatea mastita este asociată cu nivelul de lacto-ferină (r = 0,27). Există o relație pozitivă între concentrația de Lf și numărul de celule somatice din lapte, precum și între Lf și lizozimă (r = 0,35 și, respectiv, r = 0,39). Conținutul de lactoferină în timpul mastita crește de multe ori. Coeficientul de ereditabilitate pentru concentrația de lactoferină variază de la 0,15 la 0,44.
Rol important Imunoglobulinele M, A, Gi, Gj joacă un rol în protejarea ugerului de infecții. Vacile cu imunodeficiență I$G2 au o incidență crescută a mastitei piogene (purulente) Există o corelație pozitivă ridicată (r-0,9) între titrul maxim de anticorpi împotriva albuminei umane la taurii tineri și mastita clinică a surorilor vitrege. .
Au fost stabilite diferențe semnificative statistic între tipurile de BoLA și sensibilitatea la mastită la bovinele norvegiene. Antigenul BoLA W2 este asociat cu rezistență ridicată la mastită, în timp ce W16 provoacă susceptibilitate. A fost dezvăluită o legătură între alela DQ 1 MHC 11 și incidența mastitei.
Rezistența la mastită este o supratrăsătură complexă, care, la rândul său, constă dintr-un număr de supertrăsături mai puțin complexe. Această stabilitate este determinată de mecanismele de rezistență naturală. În plus, rezistența la mastită este determinată de mecanismele de apărare umorală și celulară. Datorită polietiologiei mastitei, puterea răspunsului imun la fiecare dintre numeroșii agenți patogeni este determinată de multe gene Ir.
Rezistența la mastită depinde de proprietățile morfologice și funcționale ale ugerului și de alți factori. Prin urmare, cea mai acceptabilă caracteristică a unei astfel de supertrăsături, cum ar fi rezistența la mastită, poate fi o evaluare bazată pe rata medie de incidență a familiilor și a descendenților individuali, luând în considerare markerii genetici și biochimici în viitor.
Bruceloza este o boală infecțioasă cronică a animalelor și oamenilor, cauzată de bacterii din grupul Brucella. La multe animale se manifestă ca avort, placentă reținută și tulburări de fertilitate. Boala este înregistrată în 62 de țări din întreaga lume. La unele dintre ele, infestarea animalelor este de 10-15% sau mai mare. În efectivele infectate, producția brută de animale poate fi redusă cu 15-20%. Bruceloza este de obicei transmisă la om de la animale.
Diferențele de specii și rasă. Cobai, șoarecii albi și veverițele de pământ sunt cei mai sensibili la bruceloză, în timp ce șobolanii albi, gâștele și porumbeii sunt rezistenți. Doza minimă necesară pentru infecția subcutanată a șoarecilor albi este de 5-10 celule microbiene, pentru oi - 10-15 mii, pentru vaci, porci, capre - câteva sute de mii, pentru gâște și porumbei - milioane de Brucella.
ÎN condiţii egale Zebui și bivolii sunt relativ rezistenți la bruceloză în comparație cu bovinele. În Mongolia, vitele locale sunt mai puțin infectate (3-4%) decât iacii (8-9%). Bruceloza, mastita și vibrioza pot fi răspândite în rândul raselor europene cultivate, în timp ce aceste boli nu sunt observate la animalele din rasele gri locale și rodopeane.
Oile din lână grosieră din rasa Romanov sunt mai rezistente la bruceloză decât oile din rasele Prekos și Ramboulier. La efectivele neafectate de bruceloză, incidența oilor din rasele sovietice Merino și Karabakh este mai mare, iar manifestarea clinică a bolii este mai pronunțată decât la oile din rasele Azerbaidjane Mountain Merino și Red Samukh. Oile sensibile au niveluri mai scăzute de properdină, lizozimă și un titru ridicat de aglutinine, indicând rezistență nespecifică scăzută. Aceste date indică influența eredității asupra rezistenței și susceptibilității animalelor la bruceloză.
Influența producătorilor, liniilor și familiilor. Rata de incidență a fiicelor diferiților tauri din rasa alb-negru a variat de la 8,9 la 51,9% (Tabelul 55). În medie, în fermele neafectate de bruceloză, au fost identificați aproximativ 21% dintre tații care au dat descendenți relativ rezistenți la boală, iar la 23% dintre tauri incidența fiicelor a fost mai mare de 40%. Influența producătorilor asupra incidenței bolilor la descendenți este de 8%. Nu au fost stabilite diferențe între liniile de rezistență și susceptibilitate a animalelor la bruceloză.
O evaluare a fondului genetic al familiilor a făcut posibilă detectarea familiilor rezistente, non-susceptibile, la care incidența a variat de la 0 la 61,5% (vezi Tabelul 55). Influența familiilor asupra prevalenței animalelor este de 16%.
55. Frecvența incidenței brucelozei la descendenții unor tauri și familii (conform lui V. L. Petukhov)